Los fundamentos filosóficos de la Agroecología nacen del cambio de paradigma que se ha venido dando a todo nivel en ciencia y tecnología, este cambio se da desde la perspectiva mecanicista y reduccionista imperante en la ciencia actual, hacia la visión ecológica y holística de la ciencia de contra cultura (Capra, 1998), que no es mas que el cambio de paradigma que existe entre el modelo industrialista propuesto por la Revolución Verde y el modelo holistico propuesto por la Agroecología.
En este sentido, en la Agroecología se contempla una visión de ecología moderna, que integra las corrientes de la ecología profunda de Arne Naess, de la ecología social de Murray Bookchin y del ecofeminismo de Vandana Shiva, en contraposición con el enfoque netamente naturalista de la ecología convencional.
La diferencia fundamental entre estos enfoques, radica en que la ecología convencional es antropocéntrica, es decir se centra en el hombre, ve a este por encima o aparte de la naturaleza (Capra, 1998), en contraparte, la ecología holistica, no separa a los humanos, ni a ninguna otro elemento, del entorno natural, ve al mundo no como una colección de objetos aislados, sino como una red de fenómenos fundamentalmente interconectados e interdependientes, en ella se reconoce el valor intrínseco de todos los seres vivos y se ve a los humanos como una mera hebra en la gran red de la vida (Capra, 1998).
Esta ecología reconoce como la fuente originaria de las relaciones de explotación en nuestras sociedades, la naturaleza fundamentalmente anti-ecológica de muchas de nuestras estructuras sociales, económicas y de sus tecnologías, que no son mas que expresiones del sistema de dominación occidental, que en su ultima instancia se origina en las relaciones características del patriarcado, es decir, la dominación del hombre sobre la mujer, que es el prototipo de toda dominación practicada por el ser humano (Capra, 1998).
Esta ecología al nivel de las ciencias agrícolas centra su esfuerzo en reproducir hábitats en los cuales los seres humanos se puedan vincular con su entorno mediante relaciones diferentes a las de dominación, proponiendo alternativas a los desarrollos agrícolas preponderantes, con opciones como la Permacultura, la cual tiene por objetivo diseñar sistemas orientados a la creación de entornos humanos ecológicamente sostenibles y económicamente viables.
El fundamento de este enfoque se encuentra en el entendimiento de que un ecosistema es la expresión de las relaciones entre sus componentes, por lo que cuando se combinan sus componentes (e.g., individuos, poblaciones, comunidades) en unidades funcionales mayores (e.g. ecosistema), surgen o emergen nuevas características (propiedades emergentes) que no están presentes en los niveles inferiores (Odum y Sarmiento, 1998).
De esta manera, una propiedad emergente de un nivel de organización de la materia o unidad ecológica, es aquella que resulta de la interacción funcional de sus componentes, y de este modo no puede ser predicha a partir del estudio por separado de los componentes individuales aislados o desacoplados de la unidad total (Odum y Sarmiento, 1998). Por ejemplo, cuando se combinan en cierta configuración átomos de oxigeno e hidrogeno para generar agua, esta molécula tiene propiedades diferentes a las presentes en sus átomos constitutivos por separado (Odum y Sarmiento, 1998).
Es también un ejemplo esclarecedor la relación que se da entre hongos formadores de micorrizas y las raíces de las leguminosas, en ella la combinación Hongo-Raíz es capaz de extraer nutrientes minerales del suelo de manera imposible a los dos por separado (Odum y Sarmiento, 1998).
En este orden de ideas, se puede decir que los atributos de un ecosistema son propiedades emergentes, entre ellos es particularmente importante la homeostasis, es decir las fuerzas y contra fuerzas que amortiguan las oscilaciones y que operan en todo el sistema (Odum y Sarmiento, 1998).
Actualmente es ampliamente aceptado, que la recuperación rápida después de un disturbio en un ecosistema (i.e. homeostasis), pudiese ser favorecida por la presencia de muchas especies diferentes en el paisaje (Odum y Sarmiento, 1998) (para una visión opuesta ver May, 1973).
La estabilidad del ecosistema puede medirse por medio de cuatro parámetros (Odum y Sarmiento, 1998):
1. Resiliencia: capacidad de volver por si mismo al punto inicial luego de que ha cesado una perturbación.
2. Resistencia: oposición al cambio.
3. Transiliencia: capacidad congénita de cambio.
4. Resonancia: la obtención de la misma respuesta a disturbios repetitivos.
Se cree que la estabilidad por resiliencia, es favorecida por la presencia de muchas especies distintas en el paisaje, el hecho de si una alta diversidad de especies incrementa la estabilidad por resistencia es muy debatido por los ecólogos, pero experimentos de campo realizados por Tilman et al. (1996) indican que una alta diversidad de especies en comunidades de pastizal incrementó la estabilidad del sistema durante la sequía (Odum y Sarmiento, 1998).
Relevancia en el estudio de agroecosistemas
Un agroecosistema, es un ecosistema domesticado que en muchos aspectos se encuentra en una posición intermedia entre los ecosistemas naturales como herbazales y bosques, y los ecosistemas fabricados, como las ciudades (Odum y Sarmiento, 1998).
Estos ecosistemas son accionados por la energía del sol, como los ecosistemas naturales, pero difieren de estos en algunos sentidos: existen fuentes de energía auxiliar (combustibles, trabajo animal y humano); la diversidad es reducida en gran medida por el manejo humano con el fin de masificar el rendimiento en la producción de alimento y otros bienes; los animales y plantas dominantes se seleccionan de manera artificial y no natural; y el control es externo y orientado a las metas del ser humano, en vez de interno por retroalimentación de subsistemas como en los ecosistemas naturales (Odum y Sarmiento, 1998).
Es decir que, un agroecosistema es un ecosistema que es constantemente perturbado por la acción del ser humano, la cual lo mantiene en estados iniciales de sucesión, con una biodiversidad muy disminuida, en el cual las especies que se encuentran son escogidas por el ser humano y no las producto del proceso de coevolucion, en donde los flujos de energía son determinados por el ser humano y en los cuales la redundancia trófica es casi inexistente, además este tipo de ecosistema es cíclicamente llevado a un estado de diversidad mínimo (máxima perturbación o catástrofe) al inicio de cada ciclo de cultivo (Figura 1), por lo cual se llevan a cero todas las relaciones entre los elementos del ecosistema.

Figura 1. Inicio del ciclo de siembra. Con el laboreo del suelo se llevan a cero las relaciones tróficas.
Es de resaltar que la preservación de la redundancia (la repetición de funciones), puede ser positiva, ya que esta pudiese promover la recuperación de las alteraciones producidas por las actividades humanas (Odum y Sarmiento, 1998) y es posible que sea el proceso que se encuentre detrás de la homeostasis.
Un ejemplo notable en la diferencia entre integración y redundancia de funciones de las especies dentro de su comunidad, se observa en los casos en que los insectos se convierten en plagas (Odum y Sarmiento, 1998).
La mayoría de las plagas agrícolas son especies que llevan vidas relativamente inocuas en sus hábitats nativos, pero resultan problemáticas cuando invaden un sistema agrícola (Odum y Sarmiento, 1998).
En su hábitat natural, estas especies actúan como partes de ecosistemas bien ordenados, en los cuales los excesos de reproducción y alimentación son controlados, como resultado de largos períodos de ajuste evolutivo (Odum y Sarmiento, 1998).
En situaciones nuevas (e.g., invasión de agroecosistemas), en que se carece de tales controles naturales, las poblaciones se comportan como un cáncer que puede destruir todo el sistema antes de sea posible establecer controles artificiales (Odum y Sarmiento, 1998).
Es decir, que se ha establecido una relación entre la diversidad de un ecosistema y la homeostasis del mismo, esto es un indicativo de que la falta de diversidad pudiese ser la razón de la falta de homeostasis observada en los agroecosistemas.
Estimación de la biodiversidad
Ahora bien, en este marco de ideas, la variable que deberíamos medir para comparar los ecosistemas naturales con los agroecosistemas seria la biodiversidad, sin embargo esto nos pudiese llevar a errores, ya que todas las medidas de biodiversidad con las que actualmente contamos, son producto del enfoque reduccionista de la ciencia establecida, y por lo tanto solo se centran en el estudio de los componentes por separado, y no toman en cuenta sus interacciones, lo cual se puede apreciar en el hecho de que en estos índices básicamente se toman como datos el numero de especies o clases observado (S) y el tamaño de la muestra (N), es decir ninguna variable toma en cuenta las relaciones entre las partes, solamente se toman en cuenta individualmente.
A estos estimados de biodiversidad les hacen falta tomar en cuenta la diversidad de interacciones, es decir, las diferentes relaciones entre los componentes de la comunidad, la cual no es necesariamente proporcional a la obtenida directamente de los índices clásicos de biodiversidad.
Por ejemplo, si en un ecosistema las relaciones tróficas son del tipo lineal (Figura 2), es decir el flujo de energía sigue un solo camino posible, los estimados de biodiversidad obtenidos de los índices clásicos son proporcionales a la diversidad trófica (comparar número de flechas y organismos en la Figura 2), ya que ambos se incrementan monótonamente.

Figura 2. Cadena trófica lineal.
Pero, si en el ecosistema las relaciones no son lineales (Figura 3), es decir, que el flujo de energía puede seguir diferentes rutas, los índices de biodiversidad clásicos siempre subestiman los niveles de diversidad trófica (comparar número de flechas y organismos en la Figura 3). Figura 3. Cadena trófica no lineal.
Figura 3. Cadena trófica no lineal.
Si este es el caso, con el empleo de estos índices, nunca se podría establecer el proceso responsable de la ocurrencia de homeostasis, ya que la misma es una propiedad emergente y como tal depende de las relaciones entre los elementos y no de las características individuales de estos por separado, por lo tanto, con su uso jamás se pudiesen establecer las características de la arquitectura de un agroecosistemas capaces de reproducir la autoregulación de las poblaciones de insectos, que es el fin por ejemplo de un área tan importante en la Agroecología como el manejo ecológico de plagas.
De esta manera se puede apreciar fácilmente que uno de los mayores retos que enfrentamos los agroecólogos es el construir herramientas ad hoc, que nos permitan abordar los problemas inherentes a la producción agrícola de una manera concordante con nuestras bases filosóficas.
Para elaborar un índice de biodiversidad que tome en cuenta las relaciones entre los elementos del ecosistema es indispensable utilizar las herramientas de mecánica estadística de redes complejas.
Una primera aproximación a un índice de diversidad holistico debería tomar en cuenta, como lo hacen los índices tradicionales el tamaño de la muestra (N) y el número de clases presentes en la muestra (S), tal vez relacionando estas medidas con la entropía del sistema como lo hace el índice de Shannon, pero además de esto debería tomar en cuenta el numero de enlaces en la red, el número de compartimientos o modularidad de la trama y la redundancia.
Una buena medida estándar del número de enlaces en la red es la densidad (D), esta es una medida del número de enlaces observado en la red comparados con el máximo posible (grafo completo, donde todos los nodos de la red se conectan con todos los demás), la expresión matematica de la densidad tiene la siguiente forma (Costa y Silva, 2005): Donde Vo es el número de enlaces observado y Vmax es el número de enlaces en un grafo completo con un número de vértices igual al de la red estudiada. El número de vínculos en un grafo completo con N número de nodos es (Costa y Silva, 2005):
Donde Vo es el número de enlaces observado y Vmax es el número de enlaces en un grafo completo con un número de vértices igual al de la red estudiada. El número de vínculos en un grafo completo con N número de nodos es (Costa y Silva, 2005):
 La modularidad de la trama se puede estimar mediante el estudio del mezclado de los elementos de la red, es decir, la tendencia de elementos de diferentes tipos a interactuar. Para esto se debe construir una matriz de mezclado M, compuesta de elementos mst, donde mst representa el número de vínculos que conectan elementos del tipo s con elementos del tipo t, donde los diferentes tipos representan por ejemplo gremios ecológicos.
La modularidad de la trama se puede estimar mediante el estudio del mezclado de los elementos de la red, es decir, la tendencia de elementos de diferentes tipos a interactuar. Para esto se debe construir una matriz de mezclado M, compuesta de elementos mst, donde mst representa el número de vínculos que conectan elementos del tipo s con elementos del tipo t, donde los diferentes tipos representan por ejemplo gremios ecológicos.
Esta matriz puede ser normalizada (Costa y Silva, 2005):
 Donde M representa la suma de todos los elementos en la matriz M.
Donde M representa la suma de todos los elementos en la matriz M.La probabilidad de que un elemento s tenga un enlace con un elemento t es (Costa y Silva, 2005):
 Nótese que:
Nótese que: P(s,t) y M normalizada pueden ser usados para cuantificar la tendencia de la trama trofíca a conectar nodos del mismo tipo, lo que se conoce como el mezclado (assortativity).
P(s,t) y M normalizada pueden ser usados para cuantificar la tendencia de la trama trofíca a conectar nodos del mismo tipo, lo que se conoce como el mezclado (assortativity).Se puede definir un coeficiente de mezclado como (Costa y Silva, 2005): Donde T es el número de diferentes tipos (gremios) en la red.
Donde T es el número de diferentes tipos (gremios) en la red.
Se puede apreciar que Q toma valores 0≤ Q≤1, donde Q = 1 representa una red no mezclada perfecta (los nodos de un tipo solo se relacionan con los de su mismo tipo) y Q= 0 representa una red completamente mezclada (Costa y Silva, 2005).
Esta última ecuación presenta el problema que cada gremio tiene el mismo peso, sin importar el número de especies en el gremio, una definición alternativa que evita este problema es (Costa y Silva, 2005):
Donde valores de Rc cerca de 0 indican baja modularidad de la red mientras que valores cercanos a 1 indican marcada modularidad.
La redundancia de la red puede ser estimada a través del coeficiente de agrupamiento, esta medida está relaciona con la presencia de bucles dentro de la red, por lo que representa una medida de la redundancia de la trama y de la posible presencia de retroalimentación, por lo cual esta posiblemente relacionada con la homeostasis de la red. Una definición de este índice es(Costa y Silva, 2005): Donde N∆ es el número de triángulos en la red y N3 es el número de tripletas conectadas. Un triángulo es un set de tres nodos con vínculos entre cada par de nodos, una tripleta conectada es un set de tres nodos donde cada nodo puede ser alcanzado a partir de cualquier otro nodo de la tripleta (directa o indirectamente). En la ecuación anterior el factor 3 en N se explica por que cada triangulo puede ser visto como consistente de tres tripletas. Estas medidas se pueden definir de la siguiente manera (Costa y Silva, 2005):
Donde N∆ es el número de triángulos en la red y N3 es el número de tripletas conectadas. Un triángulo es un set de tres nodos con vínculos entre cada par de nodos, una tripleta conectada es un set de tres nodos donde cada nodo puede ser alcanzado a partir de cualquier otro nodo de la tripleta (directa o indirectamente). En la ecuación anterior el factor 3 en N se explica por que cada triangulo puede ser visto como consistente de tres tripletas. Estas medidas se pueden definir de la siguiente manera (Costa y Silva, 2005):
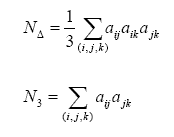 Donde la suma debe hacerse sobre todas las tripletas ordenadas (i,j,k) de los distintos nodos i,j y k.
Donde la suma debe hacerse sobre todas las tripletas ordenadas (i,j,k) de los distintos nodos i,j y k.
De esta manera la ecuación del índice de diversidad holistico tentativamente pudiese tomar la forma:
 Donde H´ es el valor del índice de Shannon. Esta aproximación toma en cuenta aspectos claves de la biodiversidad de un ecosistema, no solamente los relacionados con la riqueza y abundancia. Sin embargo para su estimación hace falta un conocimiento de las características de la comunidad muy superior al necesario para el cálculo de los índices reduccionistas.
Donde H´ es el valor del índice de Shannon. Esta aproximación toma en cuenta aspectos claves de la biodiversidad de un ecosistema, no solamente los relacionados con la riqueza y abundancia. Sin embargo para su estimación hace falta un conocimiento de las características de la comunidad muy superior al necesario para el cálculo de los índices reduccionistas.
De esta manera nos damos cuenta que en la Agroecología debido al marco conceptual con el cual se trabaja se hace indispensable generar herramientas acordes con la filosofía, tarea que no necesariamente es simple, en la cual se hace necesario el uso de las técnicas propias de la teoría de los sistemas complejos.
Esto sabemos.
Todo esta conectado
como la sangre
que une a una familia…
Lo que le acaece a la tierra,
acaece a los hijos e hijas de la tierra.
El hombre no tejió la trama de la vida;
es una mera hebra de la misma.
Lo que le haga a la trama,
se lo hace a sí mismo.
Ted Perry, 1996.
Bibliografía:
Capra, 1998. La Trama de la Vida: Una Nueva Perspectiva de los Sistemas Vivos. Anagrama.
Costa, L. da F.; Silva, F. N. 2005. Hierarchical characterization of complex networks. Journal of Statistical Physics, v. 125, p. 845-876.
May R M. 1973. Stability and complexity in model ecosystems. Princeton. NJ: Princeton. I. University Press.
Odum, E. P., Sarmiento, F. O. 1998. La Ecología: el puente entre ciencia y sociedad. Editorial. McGraw-Hill Interamericana de México.
.
.
---------------------------------------o----------------------------------
---------------------------------------o----------------------------------
.
The Apple Incident









No hay comentarios:
Publicar un comentario