Ma. Daniela Torres-Alruiz
Historia
El inicio de esta historia se remonta tal vez a 10.000 años, hasta lo que se conoce como la Revolución Neolítica, momento en el cual nuestra especie cambio sus hábitos migratorios por un comportamiento sedentario.
La razón fundamental para este cambio se debió a que descubrimos como incrementar la capacidad de carga de un ecosistema, transformándolo en un agroecosistema. Siendo esta nuestra primera experiencia en el diseño de ecosistemas.
La revolución neolítica es sin lugar a dudas la revolución tecnológica más importante que ha experimentado el ser humano. Produjo seguramente un profundo cambio en nuestra psiquis y en nuestra manera de comprender el mundo. En ese momento se modificó nuestra relación con la naturaleza, nació nuestra obsesión de dominación y tal vez las jerarquías sociales (Zerzan, 2001).
Ese camino que emprendimos, resultó en la transformación de paisajes como este (Figura 1):
 Figura 1. Ecosistema natural.
Figura 1. Ecosistema natural.En estos otros (Figuras 2 y 3):
 Figura 2. Monocultivo.
Figura 2. Monocultivo. Figura 3. Monocultivo.
Figura 3. Monocultivo.Es decir, provocó una abrumadora disminución de la biodiversidad en ciertos ecosistemas. Esta ansia de dominación hacia la naturaleza y de reducción de la biodiversidad alcanzó su punto más álgido con el surgimiento de otra revolución, la llamada: Revolución Verde.
En el marco de esta nueva revolución, el ser humano llegó a un punto tal de alienación, que desconoció todas las características y complejidades de la naturaleza, reduciéndolas a simples modelos mecanicistas.
Es por esta razón que se conoce a este tipo de agricultura como “Industrial”, no por las escalas con las que trabaja, sino por la lógica como en ella se maneja el sistema. Este esquema asume que el agroecosistema obedece a las mismas reglas que una industria. Supone que los problemas agrícolas se originan de causas similares a los que se pudiesen presentar en una línea de producción industrial.
En este marco conceptual, cualquier elemento diferente a los deseados por el gerente de la “Industria Agrícola” debe ser eliminado del sistema (Figura 4).
 Figura 4. Eliminación de la biodiversidad asociada en un monocultivo.
Figura 4. Eliminación de la biodiversidad asociada en un monocultivo.
¿Cuáles son los efectos ecológicos de este tipo de manejo del agroecosistema?
La gran mayoría de las prácticas que se hacen en el agroecosistema están relacionadas con disminuir la biodiversidad asociada e impedir que el proceso de sucesión ocurra (Figura 5).
 Figura 5. Sucesión ecológica.
Figura 5. Sucesión ecológica.
Estos modelos de producción se conocen como sistemas de sucesiones tempranas agrícolas, en ellos se presentan una serie de características:
I. La biodiversidad del ecosistema es llevado a cero cada ciclo.
II. Son favorecidas las especies invasoras (con estrategias r).
III. Los ciclos de los nutrientes se ven alterados, la biomasa no se acumula ni se cicla.
IV. El flujo de la energía no es en forma de red, sino cuasi-lineal y dirigido artificialmente.
V. Los ciclos tróficos son rotos.
El análisis de estas características es indispensable para entender el origen de las inestabilidades inherentes al monocultivo.
La primera característica hace referencia a que cuando se aplica el esquema agrícola convencional a cultivos de ciclo corto (gran mayoría de los cultivos), este involucra el laboreo intensivo del suelo al principio de cada ciclo (Figura 6). Haciendo de esta manera tabla rasa con la ya de por si escasa biodiversidad e incrementando los desequilibrios en el sistema.
 Figura 6. Laboreo del suelo. Destrucción de la biodiversidad.
Figura 6. Laboreo del suelo. Destrucción de la biodiversidad.
Sobre la segunda característica de los sistemas agrícolas de sucesiones tempranas, muy a grosso modo, y pecando de inexactitud, podemos decir que las especies que intervienen en el proceso de sucesión pueden ser divididas en dos grupos de acuerdo a sus estrategias, aquellas con estrategias tipo r, llamadas invasoras y aquellas con estrategias tipo K, llamadas dominantes (Figura 7). El resultado final de los sistemas de sucesiones tempranas es que las especies asociadas al agroecosistema (arvenses, insectos, hongos, etc.) presentan por lo general características como: alta tasa reproductiva, crecimiento rápido, buenos mecanismos de dispersión, etc. Todas estas características de las especies involucradas en los problemas de malezas, plagas y enfermedades.
 Figura 7. Estrategias de las especies involucradas en la sucesión.
Figura 7. Estrategias de las especies involucradas en la sucesión.
La tercera característica es un punto crucial, merece un Post para tratarla en detalle. Esta es una particularidad que diferencia los agroecosistemas tropicales de los de zonas templadas.
En el trópico se vive al día. Debido a las altas temperaturas características de estas zonas y a los ritmos de descomposición de materia orgánica que estas determinan, en los ecosistemas tropicales prácticamente no se acumula materia orgánica. Esta es la primera pista clave sobre la inestabilidad de los monocultivos en el trópico.
Cuando se amplía la frontera agrícola a partir de un bosque tropical, la cuenta bancaria de nutrientes esta muy próxima a cero y bajo un sistema que involucra dejar el suelo descubierto (i.e., libre de arvenses), a la merced de gotas de lluvia con grandes energías cinéticas y gran potencial erosivo, la cuenta bancaria es rápidamente llevada a cero. Este sistema sin sustento nutricional interno, se sostiene gracias a los nutrientes incorporados por el agricultor.
Esta rápida degradación de los suelos en los agroecosistemas tropicales representa una de las mayores externalidades negativas del modelo de la Revolución Verde.
Sobre las dos últimas características de los sistemas agrícolas de sucesiones tempranas centraremos esta discusión, y como se verá, ambas hacen referencia básicamente al mismo efecto del monocultivo.
El objetivo de la agricultura convencional es que el 100% de la producción del agroecosistema entre al mercado, esto es tanto como desear que, salvo las perdidas inherentes al segundo principio de la termodinámica, toda la biomasa de los frutos salga del sistema. Lo que involucra una secuencia lineal de transformación de la energía y materia desde las fuentes hasta los mercados (Figura 8). Figura 8. Cadena trófica ideal en el marco de la Revolución Verde.
Figura 8. Cadena trófica ideal en el marco de la Revolución Verde.
Este es un deseo imposible y solo sirve para generar ganancias a las empresas de pesticidas. Sin importar que esfuerzo se haga, los flujos de energía y nutrientes nunca serán completamente lineales (Figura 9). La ambición de simplificar a la fuerza la inherente complejidad de la naturaleza es otra de las causas de la inestabilidad del monocultivo, como se demostrará al final de esta discusión. Figura 9. Cadena trófica real.
Figura 9. Cadena trófica real.
Los esfuerzos por linealizar los flujos tienen el efecto negativo de romper la mayoría de la redundancia y ciclos tróficos, por lo que los monocultivos carecen de mecanismos internos de regulación poblacional. Razón por la cual son tan abundantes los problemas de explosiones poblacionales (e.g., de insectos fitófagos). Siendo esta otra externalidad negativa del modelo de la Revolución Verde.
Si el agrónomo se escapa del modelo de la Revolución Verde e intenta analizar el agroecosistema como lo que realmente es: ¡un ecosistema!. Puede apreciar que es un sistema complejo, compuesto por múltiples elementos que interactúan de diversas formas (Figura 10). Estos sistemas en muchos casos son sensibles a las condiciones iniciales, lo que significa que pequeñas modificaciones en la configuración del sistema pueden tener resultados desproporcionados. Otra característica de los sistemas complejos son las propiedades emergentes. Una propiedad emergente no depende de los elementos por separado, sino que surge de la interacción entre estos. Entre las propiedades emergentes de los ecosistemas se encuentra la estabilidad u homeostasis. La homeostasis del agroecosistema es uno de los objetivos finales de la Agroecología, alcanzarla significaría que las regulaciones de los niveles poblacionales dentro del sistema serian independientes del productor. Figura 10. Ecosistema como sistema complejo.
Figura 10. Ecosistema como sistema complejo.
Por definición, una propiedad emergente como la homeostasis no puede ser entendida mediante el estudio por separados de los componentes del sistema. El enfoque de la Revolución Verde es Mecanicista-Reduccionista, por lo que no dispone de herramientas para abordar el fenómeno de manera adecuada. Esto ha generado los grandes problemas de plagas y enfermedades de hoy en día. Estos problemas no son inherentes a la agricultura como tal, sino a un modelo erróneo de aproximación al agroecosistema. Para analizar estos sistemas se hace indispensable estudiar el todo, lo cual se puede llevar a cabo en el marco de la Teoría de los Sistemas Complejos.
Se pudiese argumentar que estos análisis y devaneos teóricos son innecesarios dado que se conocen agroecosistemas estables en el trópico, como los conucos. Esto es completamente cierto, el sistema de cultivo tipo conuco, es un sistema que presenta todos los atributos (Figura 11) que deseamos reproducir en los sistemas a mayor escala. Sin embargo, hay que tomar en cuenta, como la evidencia lo demuestra, que los conucos en cada región son diferentes. Las particularidades de cada conuco se deben a las adaptaciones del sistema de producción a las características de cada región. El ajuste del conuco a las condiciones de cada localidad se llevó a cabo de manera paulatina mediante ensayo y error.
 Figura 11. Conuco (tomada de Nuñes, 2005).
Figura 11. Conuco (tomada de Nuñes, 2005).
El problema se encuentra en que uno de los productos más nefastos de la Revolución Verde, fue homogenización en los sistemas de producción, esto trajo como consecuencia la pérdida de los conocimientos agrícolas ancestrales en vastas áreas geográficas.
El reajuste del modelo conuco a estas áreas, es un peso que no se puede dejar en manos del productor, este debe ser asumido por los centros de investigación agrícola. Si esta tarea se pretende realizar mediante bioensayos de campo, hay que tomar en cuenta que estos se deben realizar en tantas localidades como sitios se desee transformar. Lo que significa una empresa de dimensiones gigantescas.
Una alternativa es buscar en los ecosistemas naturales conocidos, las características generales de su arquitectura que les confieren estabilidad, para con base en esta información diseñar agroecosistemas que reproduzcan estos atributos. Esto se puede llevar a cabo mediante el análisis topológico de las tramas tróficas de ecosistemas naturales.
Análisis de tramas tróficas
Para estudiar las interacciones entre los componentes del ecosistema, se puede hacer uso de las herramientas de la Teoría de Sistemas Complejos, la cual es una parte fundamental (aunque muchas veces ignorada) del cuerpo filosófico de la Agroecología.
En Agroecología estos análisis tienen larga data, remontándose al trabajo clásico de Richard Levins y John H. Vandermeer (Levins y Vandermeer, 1990). Estos investigadores en un elegante articulo, demostraron con casos simplificados como la estructura de las relaciones tróficas en un agroecosistema son fundamentales en el comportamiento del sistema.
Uno de los ejemplos de Levins y Vandermeer se presenta en Figura 12A, esta figura muestra una cadena trófica lineal, compuesta por una planta (P), un insecto fitófago (F) y un insecto depredador (D). Se puede apreciar fácilmente que un incremento en la población de depredadores tendrá un efecto positivo en la de plantas al disminuir la población de fitófagos.
 Figura 12. Redes tróficas. A, red lineal. B, red no lineal.
Figura 12. Redes tróficas. A, red lineal. B, red no lineal.Levins y Vandermeer modifican ligeramente este sistema (Figura 12B), introduciendo un nuevo elemento: un insecticida (I) no específico con efectos negativos sobre F y D (rompiendo de este modo la linealidad del sistema). En este nuevo caso es interesante observar los efectos del nuevo elemento I sobre D. El insecticida I tiene un efecto directo sobre D debido a su toxicidad, pero también existe el efecto indirecto de I sobre D debido al envenenamiento de F, que es el alimento de D. Por su parte F se ve afectado negativamente por la toxicidad de I, pero esto es compensado por la disminución en la depredación de D, lo que es un efecto indirecto de la acción de I.
El uso del insecticida en este modelo incrementa las poblaciones de F. Esto no se debe a que D sea más susceptible al insecticida, sino a la estructura de la red (Levins y Vandermeer, 1990). El efecto neto de la introducción del insecticida es una disminución en la población de depredadores, generando esto lógicamente un aumento en la población de fitófagos. La moraleja de este ejemplo es que, en un sistema no lineal de solo 4 elementos, se desarrollan dinámicas que no pueden ser predichas mediante el estudio por separado de los elementos, por lo que, en el marco teórico de la Revolución Verde no se puede interpretar correctamente esta situación.
El fin último del análisis de las redes tróficas en la Agroecología es encontrar arquitecturas que generen estabilidad en la comunidad. Para realizar estos análisis hay que desplazar el foco de atención de los agroecosistemas a los ecosistemas naturales, dado que estos últimos son los sistemas que presentan la mayor homeostasis.
La diferencia fundamental, como ya se comentó, entre un agroecosistema y un ecosistema natural es la magnitud de la biodiversidad. Esto es particularmente cierto en los ecosistemas megadiversos tropicales, para hacerse una idea de la complejidad de estos ecosistemas basta observar la Figura 13.
 Figura 13. Trama trófica acuática del Parque Nacional Morrocoy, Venezuela.
Figura 13. Trama trófica acuática del Parque Nacional Morrocoy, Venezuela.
Cuando se trabaja con sistemas de estas magnitudes, los enfoques analíticos tipo Levins y Vandermeer son imposibles de realizar y se deben abordar con las herramientas de la mecánica estadística de redes complejas.
Los análisis de mecánica estadística de tramas ecológicas son un floreciente campo de estudio, que ya ha aportado interesantes resultados. Recientemente se ha sugerido (Montoya y Sole, 2003) que la arquitectura de las redes tróficas puede ser dividida estructuralmente de acuerdo a su riqueza (S) en dos clases. Aquellas que presentan riquezas bajas, se las clasifica como Poisson, en ellas todas las especies poseen un número similar de relaciones tróficas. Mientras que aquellas con riquezas altas, se las clasifica como Leyes de Potencia (también Libres de Escala), donde se encuentran pocas especies muy conectadas y la mayor parte con pocas relaciones.
Para entender esta clasificación es necesario conocer que es una distribución de grados, en el análisis de redes, el grado (k) de un nodo (elemento, especie) hace referencia al número de conexiones. En el contexto de las tramas tróficas si una especie tiene tres relaciones, entonces tiene grado 3 (k = 3). La distribución de grados es el histograma de frecuencias del número de nodos en cada grado. Que una red sea tipo Poisson, hace referencia a que su distribución de grados es Poisson (Figura 14a), una red Ley de Potencias hace referencia a que su distribución de grados es de esta forma (Figura 14b).
 Figura 14. Redes tipo Poisson y Libres de Escala.
Figura 14. Redes tipo Poisson y Libres de Escala.
Montoya y Sole (2002) estudiaron la distribución de grados de las tramas tróficas de mejor calidad conocidas (11 en total), en ellas encontraron (Figura 15) que el ajuste de las tramas tróficas de S pequeño es bueno para distribuciones tipo Poisson, pero a medida que el valor de S aumenta este ajuste se pierde. Figura 15. Distribución de grados de tramas tróficas reales. En las graficas las barras representan los valores observados y la curva el valor esperado para una distribución Poisson. (modificada a partir de Montoya y Sole, 2002).
Figura 15. Distribución de grados de tramas tróficas reales. En las graficas las barras representan los valores observados y la curva el valor esperado para una distribución Poisson. (modificada a partir de Montoya y Sole, 2002).
En la tabla 1 se puede apreciar como la distribución de grados de las diferentes tramas tróficas cambia de acuerdo con los valores de S, en esta tabla también se puede apreciar que esta tendencia se mantiene sin importar a que tipo de ambiente pertenezca la trama trófica estudiada.
Tabla 1. Características de algunas tramas tróficas Si esta es una tendencia general en la naturaleza, es decir, si la riqueza del ecosistema influye en su arquitectura, y la misma sobre sus respuestas potenciales, esto nos ofrece pistas acerca de cómo planificar la arquitectura del agroecosistema. La primera pista que este resultado nos ofrece es que, si tomamos en cuenta que un agroecosistema, por muy diverso que sea, nunca va a ser tan diverso como los ecosistemas naturales mas diversos, estos ecosistemas súper diversos no pueden ser el modelo a imitar, la escala de un agroecosistema es mas similar a la de un ecosistema natural de baja riqueza, es decir, un ecosistema con distribución de grados tipo Poisson.
Si esta es una tendencia general en la naturaleza, es decir, si la riqueza del ecosistema influye en su arquitectura, y la misma sobre sus respuestas potenciales, esto nos ofrece pistas acerca de cómo planificar la arquitectura del agroecosistema. La primera pista que este resultado nos ofrece es que, si tomamos en cuenta que un agroecosistema, por muy diverso que sea, nunca va a ser tan diverso como los ecosistemas naturales mas diversos, estos ecosistemas súper diversos no pueden ser el modelo a imitar, la escala de un agroecosistema es mas similar a la de un ecosistema natural de baja riqueza, es decir, un ecosistema con distribución de grados tipo Poisson.
Una pregunta fundamental es ¿Por qué varía la distribución de grados con el valor de S? La respuesta a esta pregunta nos pudiese dar luces sobre los aspectos claves de la topología de la red que deberíamos reproducir en el agroecosistema para hacerlo homeostático.
Para responder esta pregunta re-estudiamos la arquitectura del conjunto de redes tróficas evaluadas por Montoya y Sole. Sin embargo, a nuestro parecer 11 tramas son demasiado pocas redes para establecer a partir de ellas principios generales, para aumentar el espacio muestral generamos mediante simulación redes tróficas tipo Ley de Potencia y Poisson, en el intervalo de riquezas (S=20 a S=150) en el cual ocurre el cambio de arquitectura. Evaluamos la estructura de estas redes en busca de variaciones en su eficiencia relacionadas a la riqueza y a la arquitectura.
Para simular redes tipo Poisson se utilizó el algoritmo clásico propuesto por Erdos y Renyi (1959), el cual consiste en establecer vínculos aleatoriamente a un número dado de nodos, en este algoritmo cada extremo de un vínculo se asigna de acuerdo a una probabilidad homogénea, que en el caso de la implementación utilizada se ajustó al valor del grado promedio de las tramas tróficas conocidas.
Para simular redes Libres de Escala, se utilizó el algoritmo propuesto por Pennock et al. (2002), en el cual los nodos con mayores grados tienen mayor probabilidad de obtener nuevos vínculos, mediante el mecanismo conocido como “los ricos se hacen más ricos”. En este algoritmo se asume que en un principio cada nodo de la red tiene una probabilidad base de obtener un vínculo; subsiguientemente esta probabilidad se hace mas compleja, donde ambos extremos de cada vínculo nuevo son asignados de acuerdo a una probabilidad mixta, α para la asignación preferencial de vínculos a nodos y 1- α para la asignación uniforme, de esta manera la probabilidad de que uno de los extremos de un vínculo se una a un nodo particular es:
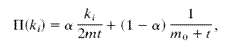 Donde mo+t es el número total de vínculos y 2mt es la conectividad en el tiempo t.
Donde mo+t es el número total de vínculos y 2mt es la conectividad en el tiempo t.
Todas las simulaciones fueron realizadas con la restricción de que el grado promedio de las redes obtenidas fuese bajo, de acuerdo a los datos empíricos que sugieren que este es el caso en las redes tróficas (Tabla 2). En este caso se restringió este valor a 13 grados, que representa el valor promedio aproximado de las redes presentadas en la Tabla 2. Las simulaciones fueron efectuadas utilizando el programa Pajek ver. 1.06 (Batagelj y Mrvar, 2003)
Tabla 2. Grados promedio (Kprom) de algunas redes estudiadas en la literatura.
A las redes obtenidas producto de estas simulaciones se les calculo el Diámetro, esta medida representa la distancia más corta entre las dos especies más distantes de la trama trófica, medida como número de vínculos (relaciones tróficas) de separación. Formalmente el diámetro es la geodésica entre los dos nodos más distantes de la red. El diámetro es una medida de la cohesión de la trama trófica.
También se realizó un análisis de vulnerabilidad, este análisis consiste en evaluar como se ven afectadas las características de las redes con la eliminación de uno de sus nodos. En este caso se estudió el número de nodos que quedan aislados en cada una de las redes producto de la remoción de un nodo, en cada caso se removió el nodo de grado mayor, es decir, las perturbaciones se hicieron dirigidas. Esta perturbación simula el efecto de eliminar completamente de un agroecosistema un elemento (e.g., una plaga)
El diámetro de la red, fue evaluado en términos del porcentaje del tamaño total de la red (Figura 16), para de esta manera estandarizar la medida y poder hacer comparaciones entre las diferentes redes. Esta variable disminuye su valor a medida que S aumenta, tanto en el caso de las redes Poisson como en el de redes Libres de Escala. Su valor es siempre mayor para redes Libres de Escala que para redes Poisson, aunque esta diferencia se hace muy pequeña para S mayores de 80. Figura 16. Porcentaje del tamaño de la red representado por el diámetro. En azul redes Poisson. En Fucsia redes Libres de Escala.
Figura 16. Porcentaje del tamaño de la red representado por el diámetro. En azul redes Poisson. En Fucsia redes Libres de Escala.
En el caso de las redes Libres de Escala, la disminución en la medida es más pronunciada, siendo la pendiente de esta disminución mayor en el caso de redes de menos de 80 nodos.
Es un hecho interesante el que la disminución en el diámetro de la red en términos de porcentaje del número total de nodos, muestre un cambio de pendiente en su taza de disminución de forma aproximadamente exponencial, observándose que la pendiente se hace muy suave para redes de S mayores de 80. Siendo este valor concordante con el cambio de arquitectura que se observa en las redes tróficas reales que han sido estudiadas.
En cuanto al número de nodos aislados luego de una perturbación (vulnerabilidad), este siempre es cero para redes Poisson, mientras que para redes Libres de Escala es muy variable (Figura 17), con una media de aproximadamente 5 especies aisladas. Figura 17. Porcentaje de nodos aislados para redes Libres de escala. En azul valor del porcentaje de nodos aislados. En fucsia promedio de la medida.
Figura 17. Porcentaje de nodos aislados para redes Libres de escala. En azul valor del porcentaje de nodos aislados. En fucsia promedio de la medida.
Nuestros resultados son concordantes y complementan a los presentados por Albert et al., (2000), quienes plantean que las redes Poisson son moderadamente sensibles a la eliminación aleatoria o dirigida de cualquiera de sus nodos, mientras que las redes Libres de Escala son muy resistentes a ataques aleatorios, esto a expensas de una alta sensibilidad a ataques dirigidos. Siendo esta una de las mayores limitaciones de la arquitectura Libre de Escala.
Esto se pudiese interpretar en términos ecológicos como que bajo la arquitectura Libre de Escala, las redes tróficas pierden cohesión cuando se elimina el nodo mas conectado, por lo que miembros de la comunidad (diferentes a la especie removida) quedan aislados del flujo de energía de la red.
Llama la atención el hecho de que si se grafica (Figura 18) la probabilidad de que por azar se elimine el nodo mas conectado, en el rango de valores de S en el cual fueron efectuadas las simulaciones, se observa un comportamiento similar al del diámetro, donde esta probabilidad disminuye de manera exponencial, con una pendiente suave para redes de S mayor a 80.
 Figura 18. Probabilidad de eliminar al nodo mas conectado de la red.
Figura 18. Probabilidad de eliminar al nodo mas conectado de la red.
La eliminación de la especie más conectada en redes Libres de Escala es un evento potencialmente peligroso para la estabilidad del sistema, se puede apreciar fácilmente en la Figura 18, que esta probabilidad es mayor para tramas tróficas de riquezas menores de 80. Por lo que la arquitectura Libre de Escala es particularmente riesgosa para redes con valores de riqueza bajos. Este comportamiento coincide con el del diámetro, el cual disminuye de manera muy suave su pendiente para S mayores a 80, valores en los cuales el diámetro de las redes bajo las dos arquitecturas son prácticamente iguales (figura 16), por lo que ninguna arquitectura es superior a la otra en términos de diámetro.
La disminución en la probabilidad de eliminar por azar al nodo mas conectado varía muy poco para redes de S mayor a 80, por lo cual no se consigue mejoras notables en términos de estabilidad aumentando los valores de S. Valor que coinciden con la evidencia empírica del cambio de arquitectura de las redes.
De esta manera nos damos cuenta que la arquitectura libre de escala para redes de S bajos es altamente riesgosa, de modo que debería ser más bien rara en sistemas naturales de estos tamaños, como es el caso observado en la evidencia empírica.
Sobre los monocultivos y los cultivos asociados
Estos resultados tienen otras interesantes implicaciones cuando se los analiza pensando en un monocultivo. Un monocultivo es un ecosistema en el cual la abrumadora mayoría de las especies están relacionas mediante algún tipo de interacción ecológica (parasitismo, depredación, mutualismo, comensalismo, competencia, etc) con una especie (i.e., el monocultivo). Esta topología se conoce en el lenguaje de redes como arquitectura Estrella (Figura 19).
 Figura 19. Red tipo Estrella.
Figura 19. Red tipo Estrella.
Bajo esta arquitectura, todas las debilidades de las redes Libres de Escala en ecosistemas de bajas riquezas se ven llevadas al máximo. La arquitectura tipo Estrella es la más inestable de todas las topologías posibles, en ella la estabilidad y cohesión del ecosistema entero depende de una sola especie. Dada esta topología, cualquier perturbación que ocurra en el ecosistema va a afectar primero al monocultivo y a través de este a todas las demás especies. En el monocultivo no existe autorregulación posible y el sistema es óptimo en la propagación de perturbaciones. Bajo esta arquitectura cualquier perturbación alcanza a todo el sistema en promedio en dos interacciones ecológicas.
Esta es la razón fundamental de todos los problemas de desequilibrios inherentes al monocultivo y es también la razón por la cual no se consigue un similar en ningún ecosistema natural. La evolución de los ecosistemas se encarga de eliminar cualquier topología que sea inestable, la tozudez del ser humano se encarga de reproducirla por doquier.
Si nuestro propósito es construir agroecosistemas estables, que reproduzcan las topologías encontradas en la naturaleza. Tomando en cuenta los niveles de diversidad posibles en un agroecosistema. Estos resultados nos indican que la manera de diseñar sistemas estables es generando arquitecturas en las cuales las apuestas no estén en una sola especie. La evidencia nos muestra que es mejor hacer innumerables apuestas, todas relacionadas entre si, donde ninguna sea indispensable. Redundancia de relaciones, es la clave de la estabilidad de estos sistemas, esta permite que las perturbaciones se compensen y modulen en una red en la cual todas las especies deben tener aproximadamente el mismo número de interconexiones.
Bibliografía:
McCann, K.S. 2000. The diversity-stability debate. Nature, 405, 228-233.
Montoya J.M. & Solé R.V. 2003. Topological properties of food webs: from real data to community assembly models. Oikos, 102 (3), 614-62.
Montoya, J.M., Solé, R.V. y Rodríguez, M.A. 2001. La arquitectura de la naturaleza: complejidad y fragilidad en redes ecológicas. Ecosistemas. Revista de Ecología y Medio Ambiente, Año X, No.2, Mayo-Agosto.
Núñez. M. A. 2005. Bases cientificas de la agricultura tropical sustentable. In Motion Magazine. June 11. http://www.inmotionmagazine.com/global/man_base.html 01/12/06, 12:00PM.
Reka, A.; Jeong, H. y Barabasi. A. L. 2000. Error and attack tolerance of complex networks. Nature 406, 378-382.
Levins, R. y Vandermeer, J. "The agroecosystem embedded in a complex ecological community" in: Carroll R.C., Vandermeer J. and Rosset P., eds., Agroecology, New York: Wiley and Sons, 1990.
Zerzan, John. 2001. Futuro primitivo y otros ensayos. Viva la República.
.
.
---------------------------------------o----------------------------------
---------------------------------------o----------------------------------
.
The Diary of Tortov Roddle: The Red Berry


 Figura 3. Cadena trófica no lineal.
Figura 3. Cadena trófica no lineal. Donde Vo es el número de enlaces observado y Vmax es el número de enlaces en un grafo completo con un número de vértices igual al de la red estudiada. El número de vínculos en un grafo completo con N número de nodos es (Costa y Silva, 2005):
Donde Vo es el número de enlaces observado y Vmax es el número de enlaces en un grafo completo con un número de vértices igual al de la red estudiada. El número de vínculos en un grafo completo con N número de nodos es (Costa y Silva, 2005):  La modularidad de la trama se puede estimar mediante el estudio del mezclado de los elementos de la red, es decir, la tendencia de elementos de diferentes tipos a interactuar. Para esto se debe construir una matriz de mezclado M, compuesta de elementos mst, donde mst representa el número de vínculos que conectan elementos del tipo s con elementos del tipo t, donde los diferentes tipos representan por ejemplo gremios ecológicos.
La modularidad de la trama se puede estimar mediante el estudio del mezclado de los elementos de la red, es decir, la tendencia de elementos de diferentes tipos a interactuar. Para esto se debe construir una matriz de mezclado M, compuesta de elementos mst, donde mst representa el número de vínculos que conectan elementos del tipo s con elementos del tipo t, donde los diferentes tipos representan por ejemplo gremios ecológicos. Donde M representa la suma de todos los elementos en la matriz M.
Donde M representa la suma de todos los elementos en la matriz M. Nótese que:
Nótese que: P(s,t) y M normalizada pueden ser usados para cuantificar la tendencia de la trama trofíca a conectar nodos del mismo tipo, lo que se conoce como el mezclado (assortativity).
P(s,t) y M normalizada pueden ser usados para cuantificar la tendencia de la trama trofíca a conectar nodos del mismo tipo, lo que se conoce como el mezclado (assortativity). Donde T es el número de diferentes tipos (gremios) en la red.
Donde T es el número de diferentes tipos (gremios) en la red.
 Donde N∆ es el número de triángulos en la red y N3 es el número de tripletas conectadas. Un triángulo es un set de tres nodos con vínculos entre cada par de nodos, una tripleta conectada es un set de tres nodos donde cada nodo puede ser alcanzado a partir de cualquier otro nodo de la tripleta (directa o indirectamente). En la ecuación anterior el factor 3 en N se explica por que cada triangulo puede ser visto como consistente de tres tripletas. Estas medidas se pueden definir de la siguiente manera (Costa y Silva, 2005):
Donde N∆ es el número de triángulos en la red y N3 es el número de tripletas conectadas. Un triángulo es un set de tres nodos con vínculos entre cada par de nodos, una tripleta conectada es un set de tres nodos donde cada nodo puede ser alcanzado a partir de cualquier otro nodo de la tripleta (directa o indirectamente). En la ecuación anterior el factor 3 en N se explica por que cada triangulo puede ser visto como consistente de tres tripletas. Estas medidas se pueden definir de la siguiente manera (Costa y Silva, 2005): 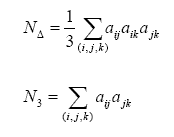 Donde la suma debe hacerse sobre todas las tripletas ordenadas (i,j,k) de los distintos nodos i,j y k.
Donde la suma debe hacerse sobre todas las tripletas ordenadas (i,j,k) de los distintos nodos i,j y k. Donde H´ es el valor del índice de Shannon. Esta aproximación toma en cuenta aspectos claves de la biodiversidad de un ecosistema, no solamente los relacionados con la riqueza y abundancia. Sin embargo para su estimación hace falta un conocimiento de las características de la comunidad muy superior al necesario para el cálculo de los índices reduccionistas.
Donde H´ es el valor del índice de Shannon. Esta aproximación toma en cuenta aspectos claves de la biodiversidad de un ecosistema, no solamente los relacionados con la riqueza y abundancia. Sin embargo para su estimación hace falta un conocimiento de las características de la comunidad muy superior al necesario para el cálculo de los índices reduccionistas.


 Por medio del análisis varianza (ANOVA) (Tabla 2) se estableció que las diferencias en el porcentaje de daño foliar en cada uno de los tratamientos (T1, T2, T3 y T4) son estadísticamente significativas (F experimental = 32.57 > F critico = 3.49).
Por medio del análisis varianza (ANOVA) (Tabla 2) se estableció que las diferencias en el porcentaje de daño foliar en cada uno de los tratamientos (T1, T2, T3 y T4) son estadísticamente significativas (F experimental = 32.57 > F critico = 3.49).  Adicionalmente con ayuda de la prueba de comparaciones múltiples de Tukey (Tabla 3 y Tabla 4) pudimos determinar que (i) el porcentaje de daño entre Tratamiento 1 y Tratamiento 3 y el Tratamiento 2 y Tratamiento 4 no presentan diferencias estadísticamente significativas y (ii) las diferencias en el porcentaje de daño entre Tratamiento 1 y Tratamiento 2; Tratamiento 1 y Tratamiento 4; Tratamiento 2 y Tratamiento 3 y Tratamiento 3 y Tratamiento 4 son estadísticamente significativas.
Adicionalmente con ayuda de la prueba de comparaciones múltiples de Tukey (Tabla 3 y Tabla 4) pudimos determinar que (i) el porcentaje de daño entre Tratamiento 1 y Tratamiento 3 y el Tratamiento 2 y Tratamiento 4 no presentan diferencias estadísticamente significativas y (ii) las diferencias en el porcentaje de daño entre Tratamiento 1 y Tratamiento 2; Tratamiento 1 y Tratamiento 4; Tratamiento 2 y Tratamiento 3 y Tratamiento 3 y Tratamiento 4 son estadísticamente significativas. 

 Señora Aniceta Guillen Landaeta (guardiana de saberes ancestrales)
Señora Aniceta Guillen Landaeta (guardiana de saberes ancestrales) Los valores de las riquezas (S) en cada una de las localidades fueron comparados por medio de un ANOVA y con la prueba de comparaciones múltiples de Tukey.
Los valores de las riquezas (S) en cada una de las localidades fueron comparados por medio de un ANOVA y con la prueba de comparaciones múltiples de Tukey.
 Las abundancias relativas para cada uno de los eventos de muestreo se presentan en la Figura 2 de cada uno de los ordenes en el Bosque y el Cultivo Agroecológico.
Las abundancias relativas para cada uno de los eventos de muestreo se presentan en la Figura 2 de cada uno de los ordenes en el Bosque y el Cultivo Agroecológico.












