.
Arbre toujours au milieu
De tout ce qui l'entoure
Arbre qui savoure
La voûte des cieux
Rainer Maria Rilke
.
.
Sesou
Apenas un adolescente, entre confundido y asustado llegué a la pequeña comunidad. Sin avisar armé mi carpa amarrillo chillón en algún sitio no bien pensado. Todavía con el entendimiento entumecido por el susto de la aventura y mientras ordenaba las pocas cosas que traía, sentí algo y volteé… para mi sorpresa, dentro de mi carpa y justo detrás de mí se encontraba un viejo indígena sentado en cuclillas. Recuerdo perfectamente la impresión que me causó la profundad de las arrugas de su rostro.
-No trajiste regalito- me preguntó. Le entregué un chocolate sin decir palabra y el igualmente se fue. Así conocí a Sesou.
Con el tiempo trabé amistad con los niños de la comunidad y después con Iona Romero, la madre de un buen número de ellos.Una amistad para toda la vida. En algún momento Iona me convenció de ir a una celebración en una comunidad cercana. Al solo llegar, vi una larga fila de indígenas que ordenadamente esperaban ser bautizados en un río por un pastor, también indígena. Junto a Wida -hija de Iona- vagabundeé al azar entre las casas de la comunidad. Al rato nos acercamos a un grupo de personas que de pie se reunían en torno a alguien. Como pudimos nos colamos y para mi sorpresa vi a Sesou, quien con un gigantesco pedazo de carne asada en la mano, gritaba agresivamente a los demás. Traté que Wida me tradujera lo que decía, pero no quiso.
-Ese es un viejo loco - me dijo- el cree que es dueño de la sabana.
Mucho tiempo después, Sesou se me acercó mientras yo holgazaneaba acostado sobre una roca.
–¿Quieres ayudar en el Conuco?- dijo y yo inmediatamente me levanté dispuesto a poner manos a la obra. Él se extrañó y se fue sin decir más. Solo después de trascurrida una semana y media partimos caminando a su lejano Conuco. Así, aprendí que hay otras formas de entender lo inmediato y de relacionarse con tiempo.
Pasamos todo el día trabajando, yo haciendo torpemente lo que Sesou me indicaba. Al inicio de la tarde paramos un rato a comer casabe con picante, le comenté que había visto algunos gusanos comiéndose las hojas los cultivos y le dije que si quería podía matarlos.
–Hay suficiente para ellos y para nosotros, todos nos beneficiamos- dijo Sesou.
También me ofrecí a apilar en un rincón los restos de troncos quemados que desordenadamente se encontraban dispersos en el conuco. Tampoco aceptó, yo simplemente pensé que era un indio terco. Al poco tiempo volvimos a trabajar, duro, muy duro hasta el atardecer. Después, de noche, acostados en chinchorros a la luz de las brasas, tuve el privilegio de escuchar, por primera vez en mi vida, el relato de cómo –en el pasado mítico- Makunaima derribó el Árbol de la Vida.
En varias ocasiones regresé a visitar a la familia Romero, a Iona y sus muchachos. Pregunté por el viejo Sesou, nadie parecía estar seguro.
- Murió hace mucho - me dijeron algunos, según otros desapareció en la sabana. Para mí fue una persona fundamental, por el decidí estudiar agricultura en la universidad. Sin embargo, ahí, ni una vez me hablaron del Conuco.
Después de graduado, un compañero de trabajo me habló maravillado de unos antiquísimos suelos amazónicos extremadamente fértiles, llamados Terra Preta do Indio, que son, sin lugar a dudas el producto de la quema de árboles en la agricultura precolombina (Petersen, Neves y Heckenberger, 2001). No pude dejar de recordar al viejo, la tarde aquella de trabajo y los troncos dispersos en el conuco. Transcurrido mucho más tiempo aun, leyendo con gran sorpresa un artículo de enigmático título en donde se describía cómo la mordida de un gusano puede ayudar a aumentar las cosechas (Poveda, Gómez y Kessler, 2010), finalmente me di cuenta… comprendí cuánto no había comprendido, lo diferente de los mundos y lo sutil que puede ser el racismo.
Tomado de: Griffon, D. 2020. Conuco: fruto del árbol Kalivirnae. En: Dinámica multifuncional de la agricultura familiar. Alimentación, ecología y economía (Ramírez y Ocampo, eds.). Colegio de Postgraduados - Universidad de Guadalajara.
Libre aquí: https://bit.ly/2L1oBFP
.
.
The Solstice
W.S. Merwin
They say the sun will come back
at midnight
after all
my one love
but we know how the minutes
fly out into
the dark trees
and vanish
like the great ʻōhiʻas and honey creepers
and we know how the weeks
walk into the
shadows at midday
at the thought of the months I reach for your hand
it is not something
one is supposed
to say
we watch the bright birds in the morning
we hope for the quiet
daytime together
the year turns into air
but we are together in the whole night
with the sun still going away
and the year
coming back
.
Constraints on selfish behavior in plants
Marina Semchenko, 2020
We are used to human behavior, and the actions of other animals, being
described as selfish, aggressive, or cooperative. Such words come up
less often when contemplating plants. Yet plants too have evolved a
fascinating array of behavioral strategies in their struggle for
resources, although these are hard to demonstrate and quantify.
Much of the world's plant biomass exists out of sight underground in the form of roots. Plants adjust how and where their roots grow according to how close neighboring—and competing—plants might be. The model extracts some of the rules about how root balls differ when grown close to neighboring plants compared with being grown in the absence of competition.
Plant roots determine carbon uptake, survivorship, and agricultural yield and represent a large proportion of the world’s vegetation carbon pool. Study of belowground competition, unlike aboveground shoot competition, is hampered by our inability to observe roots. We developed a consumer-resource model based in game theory that predicts the root density spatial distribution of individual plants and tested the model predictions in a greenhouse experiment. Plants in the experiment reacted to neighbors as predicted by the model’s evolutionary stable equilibrium, by both overinvesting in nearby roots and reducing their root foraging range. We thereby provide a theoretical foundation for belowground allocation of carbon by vegetation that reconciles seemingly contradictory experimental results such as root segregation and the tragedy of the commons in plant roots.
Ménage à Trois: Unraveling the Mechanisms Regulating Plant–Microbe–Arthropod Interactions
Gruden et al., 2020.
Plant‐microbe‐arthropod (PMA) interactions have important impacts on plant fitness, and recent studies shed light on how plants regulate responses in such complex interactions.
Biosynthetic pathways for the production of defensive and signaling compounds, and the corresponding signaling modules (mostly related to phytohormones) are key regulators both in interactions of the plant with either microbes or arthropods (two-way interactions), or when exposed to both (PMA; three-way interactions).
Most signaling modules regulating two-way interactions of plants with microbes or arthropods also operate in three-way PMA interactions, but changes in their speed or intensity (e.g., defense priming) and/or activation of additional pathways frequently occur.
These differences shape the outcome of PMA interactions and may have implications for ecologically based crop protection.

Illustration of Multiway Interactions between Plants, Microbes, and Arthropods (PMA) and the Main Signaling Pathways Orchestrating the Corresponding Plant Responses. Plants must fine-tune their molecular responses to the interaction with a plethora of organisms with different lifestyles. Microbes and arthropods interact and can alter each other’s effects on plant health through their modulation of plant responses. Continuous arrows represent the two-way interactions between the plant and the microbe or the arthropod. Discontinuous arrows represent the three-way PMA interactions. Major signaling pathways coordinating plant responses during two-way and PMA interactions are represented, namely volatile organic compounds (VOCs), jasmonic acid (JA), salicylic acid (SA), abscisic acid (ABA), and ethylene signaling (ET). Major groups of arthropod and microbe lifestyles are illustrated by particular examples, microbes on the left side of the figure, arthropods on the right. The third trophic level (indirect interaction) is also represented by parasitoids and predators of arthropods. The insert represents arthropod-associated microbes impacting the arthropod interaction with the plant. Drawing by J. Lidoy, V. Lidoy, and J. Lidoy. Abbreviations: AM fungi, arbuscular mycorrhizal fungi; PGPF, plant growth-promoting fungi; PGPR, plant growth-promoting rhizobacteria.
https://doi.org/10.1016/j.tplants.2020.07.008
.
Durable Resistance of Crops to Disease: A Darwinian Perspective
James K.M. Brown, 2015.
This review takes an evolutionary view of breeding crops for durable resistance to disease. An understanding of coevolution between hosts and parasites leads to predictors of potentially durable resistance, such as corresponding virulence having a high fitness cost to the pathogen or resistance being common in natural populations. High partial resistance can also promote durability. Whether or not resistance is actually durable, however, depends on ecological and epidemiological processes that stabilize genetic polymorphism, many of which are absent from intensive agriculture. There continues to be no biological, genetic, or economic model for durable resistance. The analogy between plant breeding and natural selection indicates that the basic requirements are genetic variation in potentially durable resistance, effective and consistent selection for resistance, and an efficient breeding process in which trials of disease resistance are integrated with other traits. Knowledge about genetics and mechanisms can support breeding for durable resistance once these fundamentals are in place.

Key features of host-parasite coevolution. (a) Reciprocal effects of interactions between host and parasite gene frequencies and fitnesses (Brown & Tellier 2011, Frank 1992, Tellier & Brown 2007). Center of diagram: A higher frequency of resistance increases the advantage of virulence, but increased virulence reduces the advantage of resistance. Top arrow: A greater cost of virulence reduces the frequency of virulence, thus increasing selection for resistance, which in turn restores the frequency of virulence. The net effect of a higher cost of virulence is to reduce the frequency of resistance. Bottom arrow: By similar logic, the net effect of a higher cost of resistance is to increase the frequency of virulence. (b) Unstable (red ) and stable (blue) polymorphisms in host and parasite gene frequencies. The unstable case is the model described in the sidebar A Simple Model of Host-Parasite Coevolution with parameters cost of resistance in host (u) = cost of virulence in parasite (b) = 0.05, cost to host of being diseased s = 0.24 and cost to parasite of incompatible interaction (c) = 1. The graph of gene frequencies spirals around and away from the unstable equilibrium point (red cross) at Req = 0.050, aeq = 0.833. The stable case has the same values of u, b, s, and c; two parasite generations per host generation; and mixed auto- and alloinfection with epidemiological parameters z = 1.4, ε = 0.1, and ϕ = 0.32 (128). The graph of gene frequencies spirals inward toward the stable equilibrium point (blue cross); the equilibrium frequency of resistance in the host is displaced slightly because of direct frequency-dependent selection on parasite. Models began with R = a = 0.02 and were run for 700 generations.
https://doi.org/10.1146/annurev-phyto-102313-045914
Agricultural diversification promotes multiple ecosystem services without compromising yield
Tamburini et al., 2020
Enhancing biodiversity in cropping systems is suggested to promote ecosystem services, thereby reducing dependency on agronomic inputs while maintaining high crop yields. We assess the impact of several diversification practices in cropping systems on above- and belowground biodiversity and ecosystem services by reviewing 98 meta-analyses and performing a second-order meta-analysis based on 5160 original studies comprising 41,946 comparisons between diversified and simplified practices. Overall, diversification enhances biodiversity, pollination, pest control, nutrient cycling, soil fertility, and water regulation without compromising crop yields. Practices targeting aboveground biodiversity boosted pest control and water regulation, while those targeting belowground biodiversity enhanced nutrient cycling, soil fertility, and water regulation. Most often, diversification practices resulted in win-win support of services and crop yields. Variability in responses and occurrence of trade-offs highlight the context dependency of outcomes. Widespread adoption of diversification practices shows promise to contribute to biodiversity conservation and food security from local to global scales.

Vote count reveals that agricultural diversification practices generally have a positive impact on biodiversity and ecosystem services. Number
of reported effect sizes with a significant positive (green), negative
(red), or neutral (gray) response to agricultural diversification,
overall (A) and to each category of diversification practice separately (B to G).

Fig. 2 Second-order
meta-analysis shows how agriculture diversification promotes
biodiversity and ecosystem services without compromising crop yield when
compared with cropping systems without these practices.(A)
All diversification practices included (324 effect sizes and 69 meta-analyses, based on 5160 original studies with 41,946 comparisons). (B)
Diversification practices targeting the aboveground environment (crop
and noncrop diversity; 91 effect sizes and 24 meta analyses). (C)
Diversification practices targeting the belowground environment
(organic amendment, reduced tillage, and inoculation; 211 effect sizes
and 55 meta-analyses). Note the difference in scale of the x
axes when comparing (A) with (B) and (C). Organic farming is included
only in the global model (A) since it often includes practices targeting
both above- and belowground environments. The number of effect sizes
and meta-analyses included in each category are displayed in
parentheses.
https://advances.sciencemag.org/content/6/45/eaba1715
1
Something startles me where I thought I was safest,
I withdraw from the still woods I loved,
I will not go now on the pastures to walk,
I will not strip the clothes from my body to meet my lover the sea,
I will not touch my flesh to the earth as to other flesh to renew me.
O how can it be that the ground itself does not sicken?
How can you be alive you growths of spring?
How can you furnish health you blood of herbs, roots, orchards, grain?
Are they not continually putting distemper'd corpses within you?
Is not every continent work'd over and over with sour dead?
Where have you disposed of their carcasses?
Those drunkards and gluttons of so many generations?
Where have you drawn off all the foul liquid and meat?
I do not see any of it upon you to-day, or perhaps I am deceiv'd,
I will run a furrow with my plough, I will press my spade through the sod and turn it up underneath,
I am sure I shall expose some of the foul meat.
2
Behold this compost! behold it well!
Perhaps every mite has once form'd part of a sick person—yet behold!
The grass of spring covers the prairies,
The bean bursts noiselessly through the mould in the garden,
The delicate spear of the onion pierces upward,
The apple-buds cluster together on the apple-branches,
The resurrection of the wheat appears with pale visage out of its graves,
The tinge awakes over the willow-tree and the mulberry-tree,
The he-birds carol mornings and evenings while the she-birds sit on their nests,
The young of poultry break through the hatch'd eggs,
The new-born of animals appear, the calf is dropt from the cow, the colt from the mare,
Out of its little hill faithfully rise the potato's dark green leaves,
Out of its hill rises the yellow maize-stalk, the lilacs bloom in the dooryards,
The summer growth is innocent and disdainful above all those strata of sour dead.
What chemistry!
That the winds are really not infectious,
That this is no cheat, this transparent green-wash of the sea which is so amorous after me,
That it is safe to allow it to lick my naked body all over with its tongues,
That it will not endanger me with the fevers that have deposited themselves in it,
That all is clean forever and forever,
That the cool drink from the well tastes so good,
That blackberries are so flavorous and juicy,
That the fruits of the apple-orchard and the orange-orchard, that melons, grapes, peaches, plums, will
none of them poison me,
That when I recline on the grass I do not catch any disease,
Though probably every spear of grass rises out of what was once a catching disease.
Now I am terrified at the Earth, it is that calm and patient,
It grows such sweet things out of such corruptions,
It turns harmless and stainless on its axis, with such endless successions of diseas'd corpses,
It distills such exquisite winds out of such infused fetor,
It renews with such unwitting looks its prodigal, annual, sumptuous crops,
It gives such divine materials to men, and accepts such leavings from them at last.
Evidence for the plant recruitment of beneficial microbes to suppress soil‐borne pathogen
Liu et al., 2020

https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.17057
.
Some theoretical notes on agrobiodiversity: spatial heterogeneity and population interactions


Genetic correlations and ecological networks shape coevolving mutualisms
Assis et al., 2020
Ecological interactions shape the evolution of multiple species traits in populations. These traits are often linked to each other through genetic correlations, affecting how each trait evolves through selection imposed by interacting partners. Here, we integrate quantitative genetics, coevolutionary theory and network science to explore how trait correlations affect the coevolution of mutualistic species not only in pairs of species but also in species‐rich networks across space. We show that genetic correlations may determine the pace of coevolutionary change, affect species abundances and fuel divergence among populations of the same species. However, this trait divergence promoted by genetic correlations is partially buffered by the nested structure of species‐rich mutualisms. Our study, therefore, highlights how coevolution and its ecological consequences may result from conflicting processes at different levels of organisation, ranging from genes to communities.
https://onlinelibrary.wiley.com/doi/10.1111/ele.13605
.
TRILLIUM
By Louise Gluck
When I woke up I was in a forest. The dark
seemed natural, the sky through the pine trees
thick with many lights.
I knew nothing; I could do nothing but see.
And as I watched, all the lights of heaven
faded to make a single thing, a fire
burning through the cool firs.
Then it wasn’t possible any longer
to stare at heaven and not be destroyed.
Are there souls that need
death’s presence, as I require protection?
I think if I speak long enough
I will answer that question, I will see
whatever they see, a ladder
reaching through the firs, whatever
calls them to exchange their lives—
Think what I understand already.
I woke up ignorant in a forest;
only a moment ago, I didn’t know my voice
if one were given me
would be so full of grief, my sentences
like cries strung together.
I didn’t even know I felt grief
until that word came, until I felt
rain streaming from me.

The origins of Amazonian landscapes: Plant cultivation, domestication and the spread of food production in tropical South America
Iriarte et al., 2020
Amazonian plant domestication is unique, differs from Eurasian processes, and needs to be study in ‘its own terms’.
Earliest colonists relied on palms, tree fruits, and underground tubers, along with terrestrial and riverine fauna.
Two areas of precocious plant domestication: sub-Andean montane forests (NW) and the shrub savannahs (SW).
First direct evidence of field polyculture agriculture within closed-canopy agroforestry and low-severity fire management.
Late Holocene spread of Amazonian anthrosols and investment in landesque capital is followed by population growth.
During the last two decades, new archaeological projects which systematically integrate a variety of plant recovery techniques, along with palaeoecology, palaeoclimate, soil science and floristic inventories, have started to transform our understanding of plant exploitation, cultivation and domestication in tropical South America. Archaeobotanical studies are providing a far greater appreciation of the role of plants in the diets of early colonists. Since ∼13ka, these diets relied mainly on palm, tree fruits, and underground tubers, along with terrestrial and riverine faunal resources. Recent evidence indicates two areas of precocious plant cultivation and domestication: the sub-Andean montane forest of NW South America and the shrub savannahs and seasonal forests of SW Amazonia. In the latter area, thousands of anthropic keystone structures represented by forest islands show a significant human footprint in Amazonia from the start of the Holocene. While radiocarbon date databases show a decline in population during the middle Holocene, important developments happened during this epoch, including the domestication of cacao, the adoption of maize and the spread of manioc across the basin. The late Holocene witnessed the domestication of rice and the development of agricultural landscapes characterised by raised fields and Amazonian Dark Earths (ADEs). Our multi-proxy analysis of 23 late Holocene ADEs and two lakes from southern Amazonia provides the first direct evidence of field polyculture agriculture including the cultivation of maize, manioc, sweet potato, squash, arrowroot and leren within closed-canopy forest, as well as enrichment with palms, limited clearing for crop cultivation, and low-severity fire management. Collectively, the evidence shows that during the late Holocene Amazonian farmers engaged in intensive agriculture marked by the cultivation of both annual and perennial crops relying on organic amendments requiring soil preparation and maintenance. Our study has broader implications for sustainable Amazonian futures.
.
Is biodiversity bad for your health?
Richard S. Ostfeld Felicia Keesing, 2017
Natural ecosystems provide services that support human well‐being, but ecosystems may also contain elements that can endanger humans. Some researchers have argued that ecosystems that support high vertebrate diversity pose a danger to human health because they are likely to support a high diversity of zoonotic pathogens, leading to the emergence of infectious diseases. We evaluated the evidence for the three necessary links in the hypothesized causal chain linking high vertebrate diversity to a high probability of emergence of infectious diseases. We found no support for one critical link—that high total diversity of vertebrate pathogens correlates with high diversity of actual or potential zoonotic pathogens. In contrast, there is now substantial evidence that high diversity protects humans against the transmission of many existing diseases. These results have substantial relevance for environmental policy.

The necessary logical steps underlying the argument that high host diversity leads to high probability of the emergence of a zoonotic disease. High diversity of vertebrate hosts must result in high total diversity of pathogens within the vertebrate community, which in turn must lead to high diversity of actual or potential zoonotic pathogens (those that can infect humans and cause disease), which in turn must increase the probability of new emergence events. Although a link between host diversity and parasite diversity is relatively well established, effect of host diversity on viral and bacterial pathogens (arrow 1) is not. Evidence does not support a link between overall pathogen diversity and that of actual or potential zoonotic pathogens (arrow 2). Some evidence supports correlations between diversity of zoonotic pathogens and the likelihood of zoonotic emergence (arrow 3), but with important caveats described in the text.
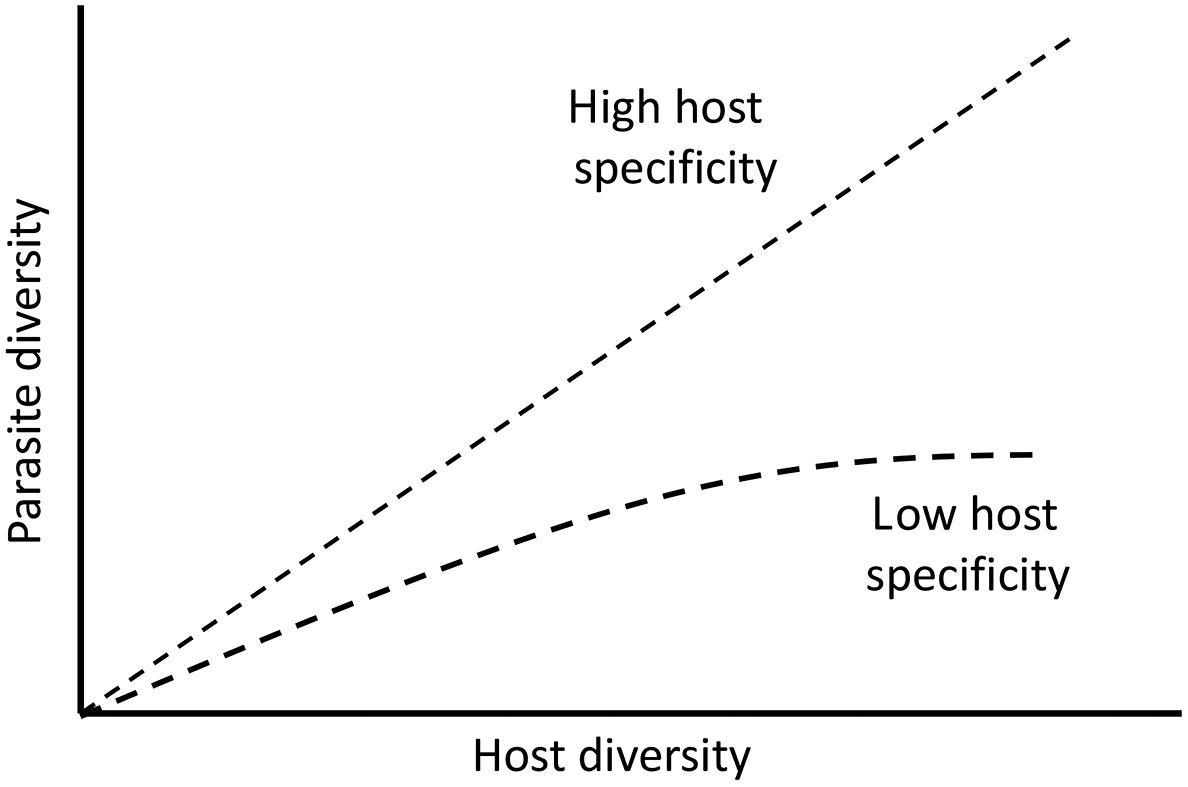
Schematic diagram of how parasite diversity is expected to vary with host diversity when parasites show high host specificity (upper curve) and when they show low host specificity (lower curve). In the latter case, the sharing of parasites between hosts means that the diversity of parasites will saturate as host diversity increases, resulting in little or no additional increases in parasite species at high levels of host diversity.

Schematic representation of typical rank–abundance curves, in which the relative abundance of each species is represented on the vertical axis and the rank of each species, from highest to lowest abundance, is given on the horizontal axis. Contrasted are two scenarios, a relatively low‐diversity community in blue and a relatively high‐diversity community in green. The curves represent the common observation that higher‐diversity communities include more species that are rare and fewer that are common. The species added (right‐hand orange circle) in higher‐diversity communities are not likely to be the sources of zoonotic pathogens, whereas the most abundant species in lower‐diversity communities (left‐hand orange circle) are often the sources of zoonotic infection.
.
















